|
3. エタノール存在下における増殖の影響10,11) そのような醪中の酵母のリン脂質脂肪酸組成は,酒母のそれに比べて,パルミチン酸(16:0)含量がさらに増大し,速醸もとを酒母とする清酒醪では,これにともなってリノール酸(18:2)含量が減少していた.そのために,いずれの酒母を用いた仕込でも不飽和度((% monoene+2[% diene])/100)12)が低下したが,生もと仕込の醪と速醸もと仕込の醪の間には,リノール酸(18:2)含量の違いによる不飽和度の差が,依然として認められた. そこでつぎに,このような醪中でのリン脂質脂肪酸組成の変化に,エタノール存在下に増殖することが及ぼす影響について検討した. 4%,または8%濃度のエタノールを含む培地で,パルミチン酸(16:0),またはリノール酸(18:2)を添加して酵母を培養し,リン脂質脂肪酸組成をエタノールを含まない培養によって得られた菌体と比較した(Table 5).
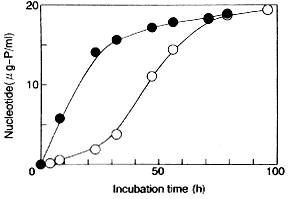
エタノール存在下で増殖するとき,培地中にパルミチン酸(16:0)が添加されていると,リン脂質中のパルミチン酸(16:0)が著しく増大するとともに,ミリストオレイン酸(14:1),パルミトオレイン酸(16:1)およびオレイン酸(18:1)が減少した.他方,エタノール存在下にリノール酸(18:2)が添加された場合には,オレイン酸(18:1)の増大とこれに照応したパルミチン酸(16:0),ミリストオレイン酸(14:1),パルミトオレイン酸(16:1)の減少が認められ,リン脂質の不飽和度は増大した.このように,パルミチン酸(16:0)とリノール酸(18:2)添加の間には,エタノールによるリン脂質脂肪酸の組成の変化に違いが認められた.また,エタノールのない環境で増殖した菌体を12%エタノールを含む培地に移して置換培養を行っても,その脂肪酸組成は変わらず,脂肪酸組成変化は増殖にともなう適応と考えられた.これらのことから,清酒醪中で,酵母リン脂質アシル基にパルミチン酸(16:0)が増大するのは,段仕込によってエタノール存在下に増殖する際にパルミチン酸(16:0)を利用した結果と考えられた. Saccharomyces cerevisiae NCYC431が,エタノール存在下に増殖するとき,オレイン酸(18:1)が増大することが報告されており,13)このことから,不飽和度の増大がエタノールに対する適応であると考察されている.しかしながら,パルミチン酸(16:0)添加培養菌体を18%エタノールを含むGYP培地に懸濁したときの15℃における死滅速度定数(d-1)は,培養時のエタノール濃度が0,4, 8%のときそれぞれ0.328, 0.269, 0.142となり,リノール酸(18:2)添加培養菌体のそれはそれぞれ,0.506, 0.453,0.161と小さくなっていったが,いずれの場合もパルミチン酸(16:0)添加培養酵母が高い生存率を示した.これらのことから,エタノールを含む培地中で増大した菌体のエタノール耐性の増大を,菌体のリン脂質不飽和脂肪酸の増大の結果として,一義的に結び付けられないことを示している. また,これらの菌体を20%エタノールに懸濁し,約10時間にわたるヌクレオチド漏出経過から,膜透過係数を算出して比較した(Table 6).
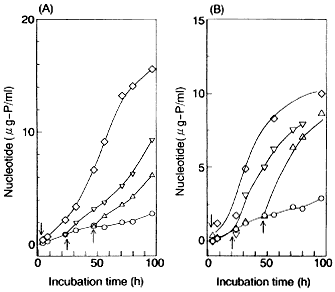
いずれの脂肪酸添加においても,培養時のエタノール濃度が上がるにつれて,膜透過係数は小さくなったが,それぞれの培養条件において,パルミチン酸(16:0)添加培養酵母の方が小さな値を示した.これらのことから,エタノールに対する膜のバリアー能は,リン脂質アシル基の鎖長と飽和度に依存した疎水結合力に関係していると考えられた. また,4%および8%エタノール存在下に培養された菌体の脂肪酸組成に大きな相違がないにも関わらず,8%エタノール存在下に培養した菌体の膜透過係数がさらに小さくなったが,これは20%エタノールに懸濁した初期にのみ観察された(Fig.7).
初期に見られるこのようなヌクレオチドの漏出抑制は,グルコース,フラクトース,マンノースのような発酵性糖を添加することにより継続して観察されたが,非発酵性糖の添加は無効であることから,解糖系によって生成されるATPに依存していると考えられた. そこで,50mMグルコースを加えた20%エタノールに菌体を懸濁してヌクレオチド漏出を抑えておき,モノヨードアセトアミドの添加効果を見た(Fig. 8A).
解糖系を阻害するモノヨードアセトアミドの添加に伴い,ヌクレオチドの漏出速度が増大し,膜透過性が大きくなると考えられた.細胞膜ATPaseの特異的阻害剤であるスティルベストロールの添加によっても同様に,添加に伴いヌクレオチドの漏出速度が増大した(Fig. 8B).このことは,膜ATPase活性によって実現される細胞内のイオン恒常性が,膜構造の安定性に寄与していることを強く示唆するものと考えられる.14) |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
[おいしさの秘密は生もとづくり]へ戻る |
お酒は20歳になってから。 お酒はおいしく適量を。
Copyright(C)Kiku-Masamune Sake Brewing Co., Ltd.
1999-2003 All rights reserved.